Uncategorized Tuesday, 2024/03/19
Hainan Medical College's Ren Ruibao, Cai Wangwei, and Shanghai Jiao Tong University's Chen Zhu jointly published a research paper on PNAS titled "The impact of age and number of mutations on the size of clonal hematopoiesis", which reported an in-depth analysis of Clonal Hematopoiesis (CH) in the elderly population with longevity (age ≥ 90 years old) and general health (age 60-89 years old). This study utilized targeted deep sequencing and found that the occurrence and development of CH are closely related to the expression of age and aging biomarkers.
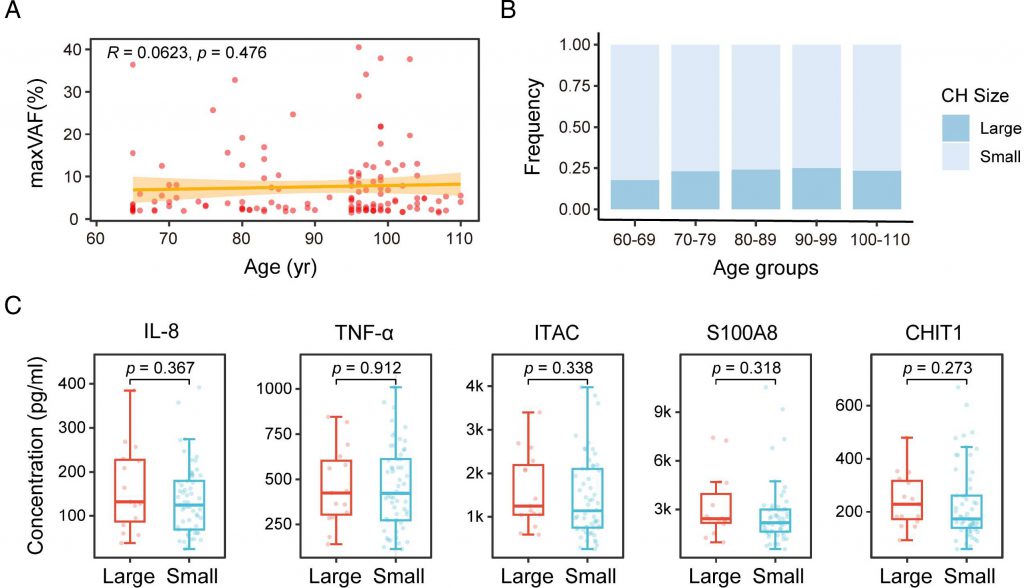
The incidence of CH in the elderly group with a long lifespan is significantly increased, and the frequency of TET2 and ASXL1 gene mutations is significantly increased, indicating that a certain degree of CH may be beneficial for prolonging lifespan. Interestingly, the size of CH is not significantly correlated with age (60-110 years old), nor is it significantly correlated with the expression of aging biomarkers. On the contrary, there is a strong correlation between the size of large CH and the number of mutations in each individual. These findings provide a biomarker for risk assessment of CH, and also indicate that the evolution of CH is influenced not only by age but also by other factors.
Our Featured Products
Hematopoietic stem cells (HSCs) gradually lose their proliferative potential during the aging process. Meanwhile, during the process of self-renewal, HSCs continuously accumulate DNA mutations. Certain mutations in HSCs can cause increased proliferation and abnormal differentiation. The clonal expansion of hematopoietic stem cells and their offspring driven by somatic mutations is called clonal hematopoiesis (CH). As age increases, CH becomes increasingly common, known as age-related clone hematopoiesis (ARCH). The presence of somatic mutations in malignant tumor-related genes in the blood, without evidence of hematological malignancy, developmental abnormalities, or decreased blood cells, and with a variant allele frequency (VAF) ≥ 2%, is called clonal hematopoiesis of uncertain potential (CHIP).
Previous studies have shown that the prevalence of CHIP steadily increases with age. Common CHIP mutations affect multiple genes, such as DNMT3A, TET2, PPM1D, TP53, STAT3, IDH1, IDH2, and ASXL1, most of which are known to be associated with hematological malignancies. These mutated genes can be classified as epigenetic regulatory factors, splicing factors, DNA damage response factors, and hematopoietic transcription factors. In addition to age, the occurrence and development of CHIP are also influenced by factors such as inflammation, cancer-related treatments, smoking, and genetic factors.
CHIP is associated with an increased risk of hematological malignancies such as acute myeloid leukemia (AML), unexplained clonal leukopenia, myelodysplastic syndrome, and non-hematological diseases such as cardiovascular disease, solid tumors, and autoimmune diseases, as well as an overall increased mortality rate. CHIP can also trigger inflammatory responses in the body and exhibit strong tolerance to this inflammatory environment, enhancing the adaptability of CH and increasing the risk of sepsis, severe coronavirus infection (COVID-19), and other infections.
Not all CH will evolve into hematological malignancies or other diseases. Therefore, risk assessment of CH is crucial in disease prevention and clinical practice. Research has shown that the evolution of CH is influenced by both mutation types and non-mutation-related factors. The study also found that the size of CH is associated with disease progression and prognosis, and larger clones are associated with higher disease risk. Previous studies have found that individuals with high VAF (≥ 10%) have a significantly increased risk of developing lung cancer, gout, and major adverse cardiac events (MACE). The size of CH is also included in the CH risk score for predicting myeloid malignancies. However, the factors that affect CH size and their potential molecular mechanisms are still unclear.
Somatic mutation-driven clone amplification occurs not only in the hematopoietic system, but also in normal tissues throughout the body. Although certain clonal expansions can evolve into cancer and/or promote the development of other diseases, certain clonal expansions may also exert beneficial homeostatic effects by combating stress and aging conditions in certain states. As age increases, the hematopoietic capacity of HSCs gradually decreases, and this change becomes more pronounced after the age of 70. Although CH has been detected in various age groups, systematic research has not yet been conducted in long-lived populations (90 years and above).
Based on this, the study compared the prevalence of CH between the long-lived (age ≥ 90 years) and ordinary (age 60-89 years) elderly groups, and found that the prevalence of CH in the long-lived elderly group significantly increased, with a significant increase in TET2 and ASXL1 mutations. Surprisingly, within the age range of 60 to 110 years, there was no significant correlation between CH size and age, nor was there a significant correlation with the expression of aging markers. On the contrary, there is a significant correlation between the size of CH and the number of mutations in each individual. These findings provide biomarkers for risk assessment of CH and indicate that the evolution of CH is influenced not only by age but also by other factors.
Related Products and Services
Cytokines Cancer Drug Targets Immune Checkpoint Proteins Protein Interaction Service Protein Expression and Purification Services Drug Discovery Screening Protein Pathway Profiling
Reference Wang K, Zhang W, Yi L, Zhao M, Li PY, Fu MH, Lin R, Zhu YM, Li JF, Yang WP, Fang H, Chen Z, Cai WW, Ren RB. The impact of age and number of mutations on the size of clonal hematopoiesis. Proc Natl Acad Sci USA. 2024 Feb 20;121(8):e2319364121. doi: 10.1073/pnas.2319364121. Epub 2024 Feb 15. PMID: 38359296; PMCID: PMC10895265.